
RESEARCH
Metazoans have organs with well-defined shapes and sizes that are specific to their species. This requires mechanisms that regulate the number, sizes and types of cells within developing organs. When these mechanisms fail, individuals suffer from disorders and malformations, such as microcephaly, spina bifida, cancer and others. Prior studies have identified many molecular components that are involved in growth and proliferation. However, it is still unclear how cellular behaviors are controlled and coordinated at the level of an entire organ.
We are interested in the mechanisms that control tissue growth and pattern formation and ensure reproducible developmental outcomes. We study these mechanisms in the developing vertebrate spinal cord, focusing on mouse and chick.
An essential part of the vertebrate central nervous system, the spinal cord relays motor and sensory information between the body and the brain. In this organ, a highly conserved pattern of neural progenitor subtypes is established along the dorsoventral (DV) axis as the tissue grows, which makes it an ideal system to study morphogen-mediated development. It is an experimentally tractable system, where embryonic stem cell differentiation, the mouse and chick model systems can be used in a complementary manner. We and others have developed tools that allow quantitative measurements and analysis of spinal cord development. The questions about size, reproducibility, patterns and dynamics that we ask are inherently quantitative, hence in our work we aim to obtain quantitative and dynamic data. To combine these data with mathematical modeling and theoretical descriptions, we have a mix of experimentalists and theorists in the group and we work in close collaboration with biophysicists.
Morphogen gradients and pattern formation
To achieve defined sizes, organs must balance rates of cell division, cell loss, and the proportions of cell types produced. A common mechanism that controls both cell number and cell type composition is the formation and interpretation of morphogen gradients. Morphogens are conserved signaling molecules that emanate from localized sources and spread to form spatial gradients within target tissues. Such gradients provide growth and patterning cues that determine cell number, fate, and positioning (see review).
How do morphogen signaling gradients form in the neural tube? What is their robustness and reproducibility, and how is the spatiotemporal dynamics of signaling regulated? To address these questions, we previously measured the Shh and BMP signaling gradients during the first three days of neural tube development and quantified their variability (Kicheva et al 2014, Zagorski et al 2017). We showed that in the neural tube, cells integrate information from the opposing morphogen gradients of Shh and BMP and this maximizes the precision of pattern formation (Zagorski et al 2017).
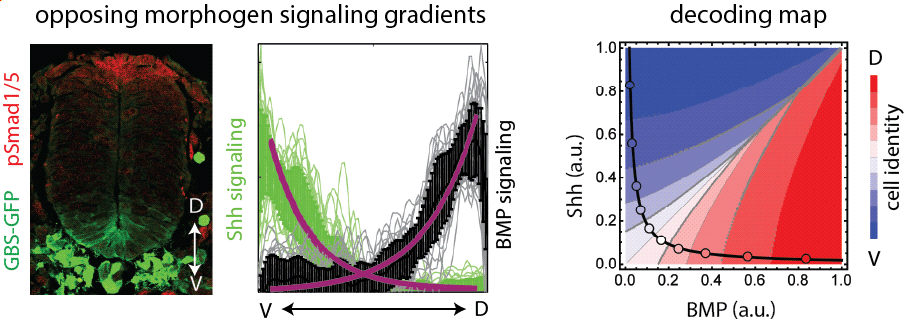
Following up on this work, we investigated what mechanisms contribute to the dynamics of both dorsal and ventral signaling gradients in the neural tube. To this end, we established a 2D spinal cord organoids that form self-organized patterns characteristic of the dorsal neural tube (Lehr et al 2024). This system is ideal to study both tissue growth and pattern formation in an accessible and quantitative manner. Using this system, we found that self-organized pattern formation depends on a BMP morphogen signaling gradient with specific biphasic temporal dynamics (Lehr et al 2023). In collaboration with biophysicists, we developed a data-driven theoretical framework and showed that the biphasic BMP dynamics are produced by a temporal relay mechanism that couples fast inhibition and slow positive regulation of BMP signaling.

These studies opened many new questions that we are currently pursuing in the lab. We are investigating the spatial dynamics of morphogen signaling in organoids and in vivo, the cross-talk between signaling pathways, the downstream interpretation of morphogen gradients in pattern formation and growth control. We are specifically interested in the feedbacks between morphogen gradients and tissue growth.
In parallel with our in vitro work, we are continuously working with embryos. Using a biophysical model coupled with experimental data, we recently showed that after its initial specification, the Shh morphogen source expands by tissue growth, thereby coupling the gradient shape to tissue size (Ho et al 2024).
Size and shape of the neural tube
The early stages of neural tube development are remarkable in that they involve rapid growth and profound morphogenetic changes in which the neural plate bends closes to form a neural tube. We found that during these early stages, the neural epithelium is in a fluid-like state and undergoes substantial cell rearrangements (Bocanegra et al, 2023). We also found that the declining growth rate over time leads to a decrease in the cell rearrangements. These changes in growth rate also affect the anisotropy of tissue growth (Guerrero et al, 2019) and thereby the shape of the tissue.
In our current work, we are investigating what are the mechanisms that control the growth rate, what is the role of morphogen signalling as well as mechanical forces in growth control.

Grants
ERC Consolidator Grant (2022)
Mechanisms of tissue size regulation in spinal cord development

SFB Grant, FWF Austria (2020, 2024)
Stem cell modulation in neural development and regeneration
ERC Starting Grant (2015)
Coordination of patterning and growth in the spinal cord
